
Quick reminder: Applications for the J.E. Weaver grants ($1500) for graduate students working on conservation topics related to the Great Plains are due March 3, 2023. Information can be found here.
This one’s for the real prairie nerds out there. It’s long, full of data, and will probably appeal mostly to people who restore prairies or study prairie restoration. Apologies to those of you who mainly follow this blog for the pretty photos…
Despite the obscene number of graphs, I do think there’s a pretty fascinating story here. I’ll do my best to tell it in a compelling way. The story starts with the pattern with which a brand new prairie plant community assembles itself. That’s a good tale in itself, but then I compare that pattern to two other nearby plantings with surprising (to me) results. Finally, the significant role of land stewardship and its impact on establishment fills the third and final chapter of the story.

First, the basic information:
The 100 acre Derr Pivot prairie restoration was planted into a former soybean field in the fall of 2001. We call it a 2002 restoration because 2002 was its first growing season. The site is located in some sandy hills south of the river at The Nature Conservancy’s Platte River Prairies. Seed for the project was locally-harvested and the seed mix included 165 plant species. The first three growing seasons for the planting were drought years but it established well with few invasive species issues other than some Siberian elm trees.
Since its third growing season (2004), I’ve been collecting plant community data from the site. Every other year, I go out with a 1x1m plot frame and list the plant species within 70-100 sampling plots scattered across the site in a stratified-random pattern (not permanent plots). From that, I can calculate both the species richness (number of species) and floristic quality of each square meter plot. Averaging those across the site gives me the mean species richness and mean floristic quality for that year.
(Reminder, if you want better views of graphs and photos, click on them. If you’re reading this within an email message, click on the post’s title to open it on the website and then clicking on images will work for you.)
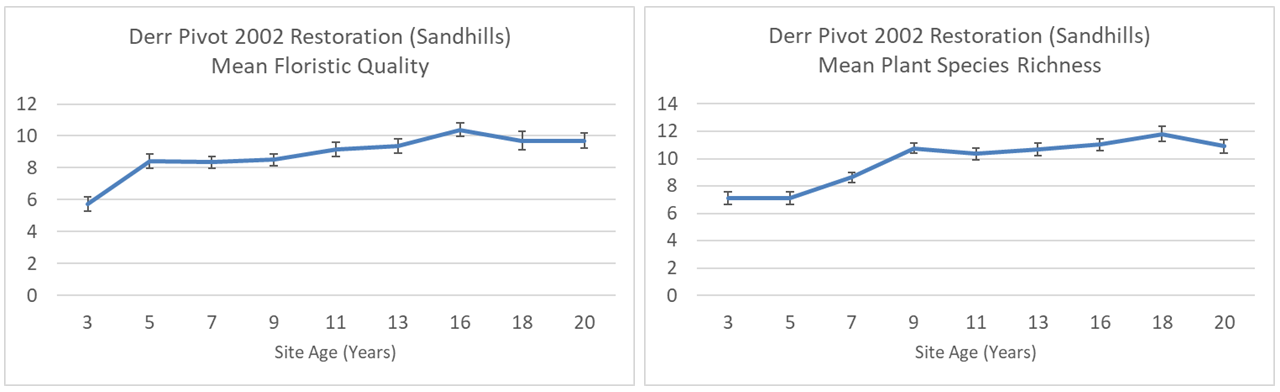
As you can see from the two graphs above, both the mean species richness and mean floristic quality increased during the first 8-10 years or so and then have leveled off. I’ve also found 143 plant species in those surveys, though that’s not a complete inventory of species, since those 1×1 plots don’t sample a very big percentage of the area each year.
That’s the boring background information. If you’re still with me, here’s where I’ll try to make it more interesting.
.
CHAPTER 1 – A PLANT COMMUNITY FORMS
In addition to calculating richness and floristic quality metrics, I can also look at the frequency of occurrence of each species (the percent of 1×1 plots each species was found in) through the years. It doesn’t work well for plant species of low abundance, but for major grasses and forbs, I can look at how the abundance (essentially) of each species changed over time. Most of those species increased in occurrence over time and then leveled off. Others, like the three grasses and one sedge in the graph below, appear not to have hit their peak yet, even after 20 years.
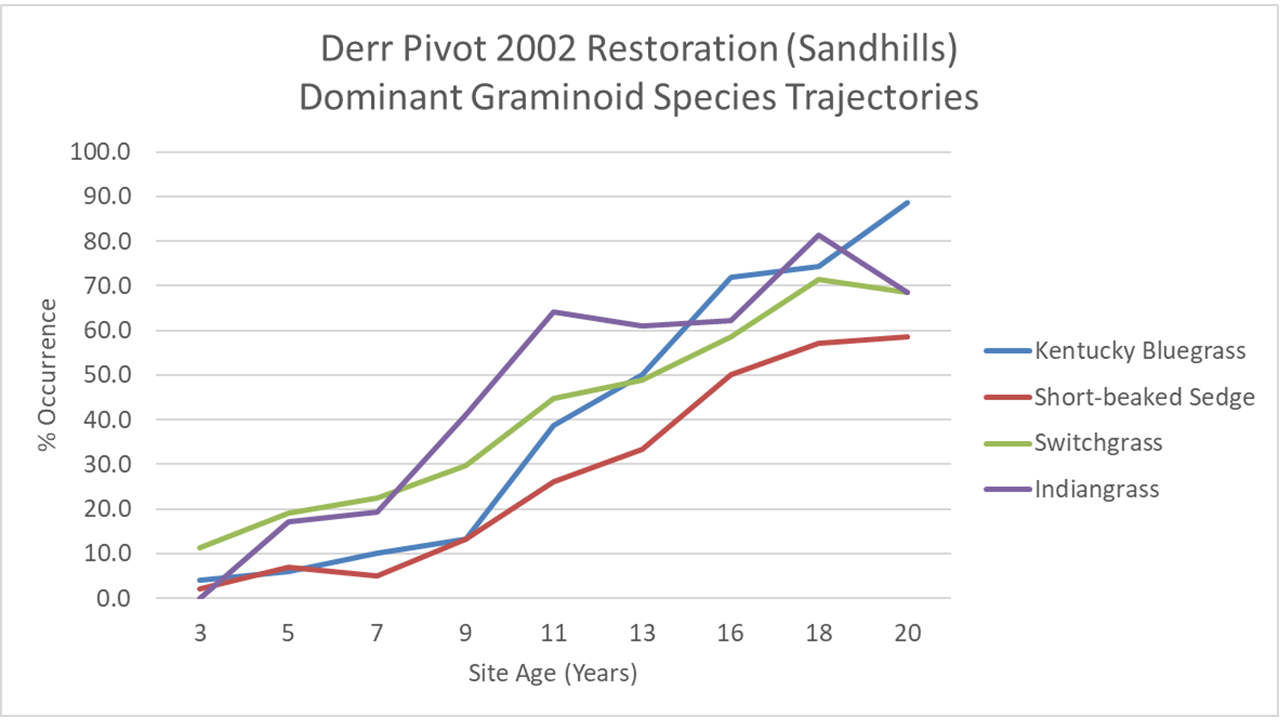
The graph shows the increasing frequency of occurrence of three grasses and one sedge (Carex brevior) over the first 20 years of this restored prairie’s life. Kentucky bluegrass wasn’t seeded, of course, and short-beaked sedge had a limited amount of seed, which helps explain why both started slowly. However, we had lots of switchgrass and Indiangrass seed, so why is it taking so long to fill in? That’s especially interesting because big bluestem – shown a few graphs below this – jumped out of the ground and was ubiquitous within a handful of years.
These graphs illustrate the assembly of a brand new prairie plant community over a 20 year period. Some species showed up in abundance within the first few years, but most started slowly and spread over time. The rate at which species increased in abundance to a point at which they seemed to reach their saturation point varied quite a bit.
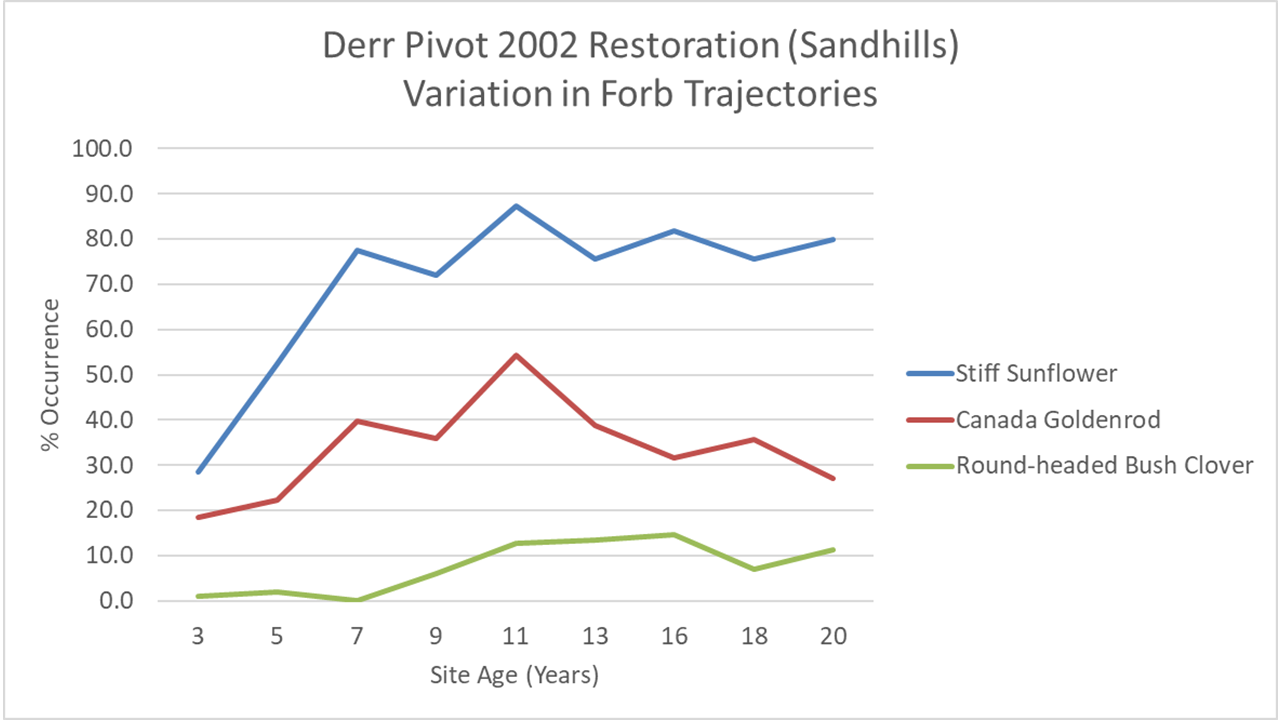
The graph below, for example, shows three very different patterns. Stiff sunflower (Helianthus pauciflorus) raced out to a fast start, and by year seven I was picking it up in 4 out 5 sampling plots. It has stayed roughtly at that level ever since. Canada goldenrod (Solidago canadensis) increased steadily until it was in half of my sampling plots in the 11th year of the planting, but then decreased just as steadily back to only 20% occurrence by year 20 – the same place it had been in year three. Round-headed bushclover (Lespedeza capitata), which is mostly found on hill tops and higher slopes, also took about 11 years to reach its peak, but, unlike the goldenrod, has maintained that level of abundance ever since.
I mentioned big bluestem earlier. The graph below shows that big bluestem was already above 50% occurrence in the third year of the planting. That’s a significant difference from switchgrass, which was at 11% that year, and Indiangrass, which didn’t show up in any of my sampling plots at that stage. I don’t have an explanation for that, especially because harvested the grass seed myself with an old Massey combine, and while big bluestem was the most common of those three species in the sites I harvested, there was lots of Indiangrass and switchgrass too.

Even more fascinating to me, though, is the contrasting pattern of occurrence between big bluestem and Canada wildrye (Elymus canadensis). We worked hard to harvest lots of Canada wildrye seed because we know it tends to establish quickly but isn’t competitive against the other species we plant. That certainly held true here. It was at 86% occurrence at year three. But then, as big bluestem increased in abundance, Canada wildrye decreased, leveling out at 40-50% frequency of occurrence by year 9. It has held steady there since. I love being able to see evidence that supports the way I thought these two species worked in our plant communities. Big bluestem is a dominant force and Canada wildrye is more opportunistic, though both are perennial species.
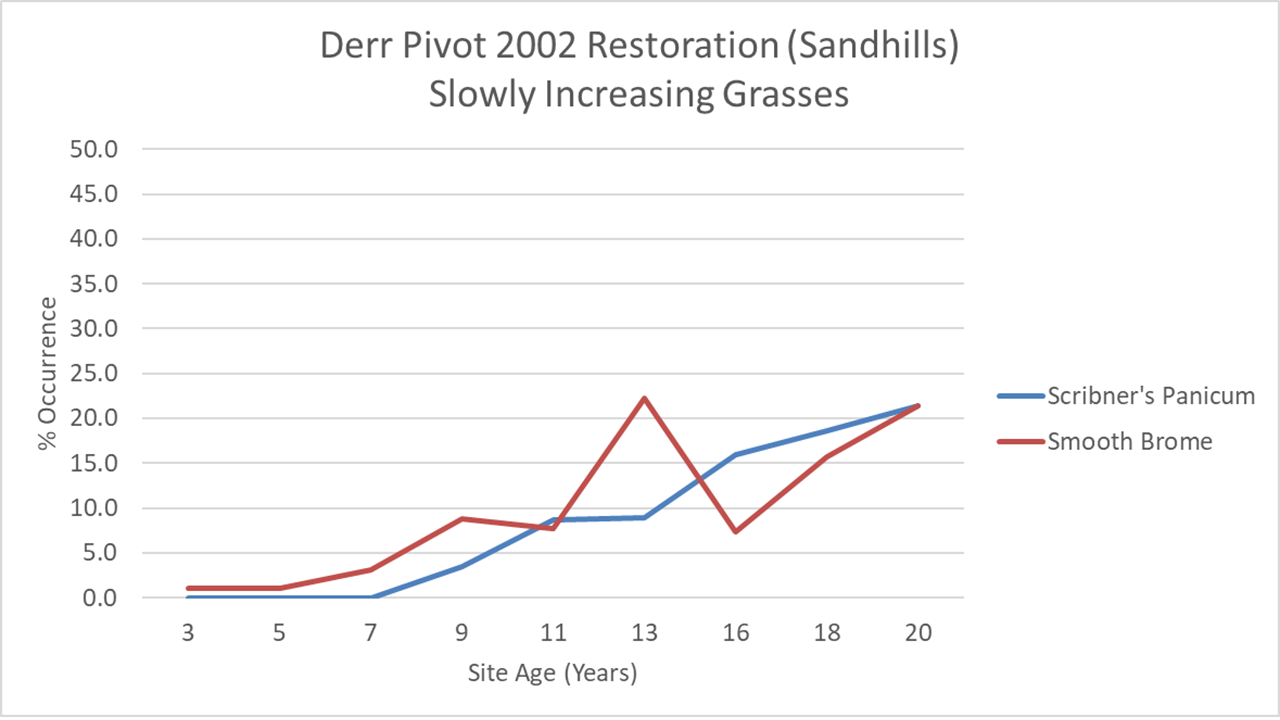
In contrast to big bluestem and Canada wildrye, two other grasses started out very slowly, and continue to increase very gradually. Scribner’s panicum (Panicum oligosanthes) is a great little native grass for which we had harvested very little seed. It’s not surprising, then, that it took a while to get going, and I’m encouraged that it continues to become more and more a part of the community. Smooth brome, on the other hand, can go jump in a lake as far as I’m concerned, but seems intent on slowly creeping in from the edges of the site. That could become a more significant issue over time if it suppresses plant diversity. So far, though, even at sites where it has already become much more widespread, our management seems to be keeping it from having a big effect on the community. We’ll see.
Finally, I wanted to talk about two grasses that are tied mostly to hill tops where the soil texture is most sandy. That restrictive habitat preference means they’ll be limited to a lower level of abundance than other species and both appear to have reached it. Junegrass (Koeleria macrantha), however, hit its plateau in year 5, while it took prairie sandreed (Calamovilfa longifolia) about 13 years to get to the same place (see graph below). I wouldn’t have guessed that, based on our seed harvest. We collected Junegrass by hand and didn’t have a particularly bounteous harvest, as I recall. I harvested prairie sandreed with our combine, and worked hard to get a lot of it, knowing it would do well in the sandhill habitat we were planting.
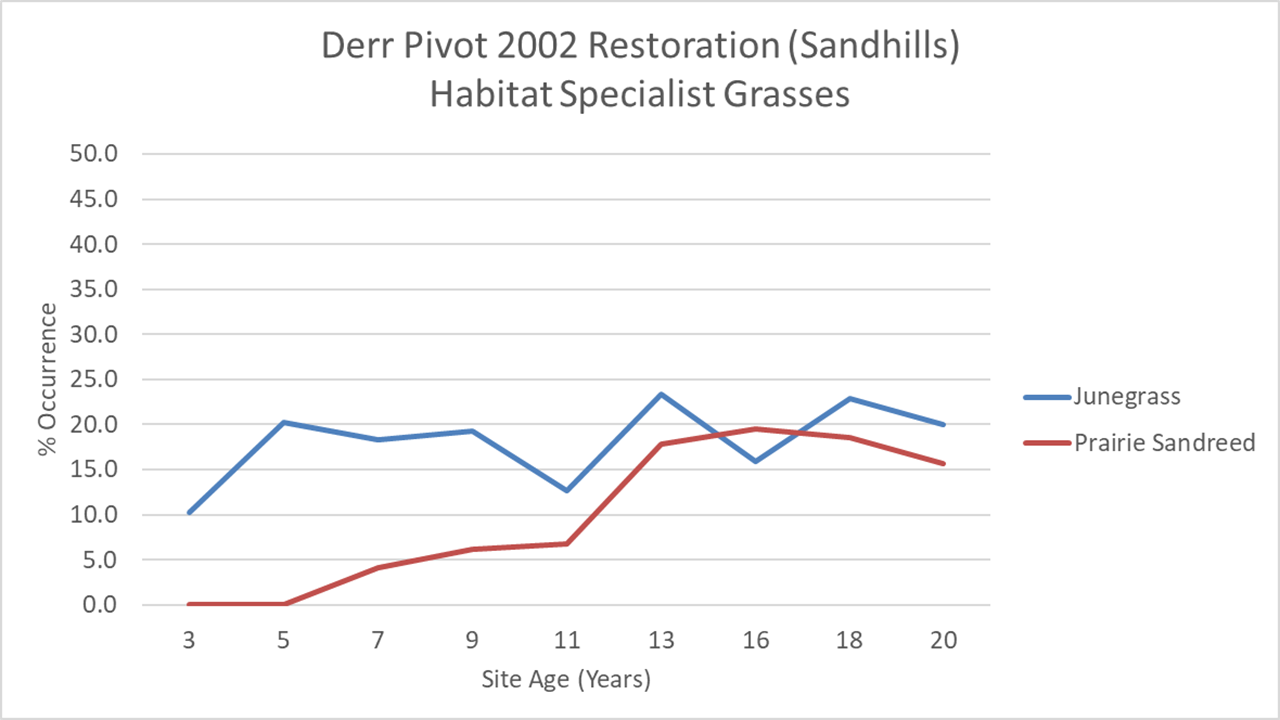
I think the explanation for Junegrass’ rapid rise is that it spread quickly by seed from its initial establishment points. It showed up right away from the seed we planted and then filled in the space around those first plants very quickly. Prairie sandreed, on the other hand, seemed to take a while to become full-size blooming plants, and appears to have spread primarily through rhizomes. Sandreed clones are now getting large (but are diffuse, not monocultures) and many are bigger than my kitchen. (You know my kitchen, right? The standard by which ecologists measure grass clones.)

.
Ok, that’s Chapter 1. Compelling? No? Ok, give me a chance with Chapter 2 before you bail.
CHAPTER 2 – EVERY SITE IS DIFFERENT. MAYBE?
Having mined the data from the Derr Pivot site, I decided it would be interesting to compare the patterns of species establishment to that of other nearby plantings we’d done. I expected to see a lot of variation in those patterns because anyone who has done restoration work knows there’s a lot of unpredictability in that process. As it turned out, I found much more similarity than I’d expected.
I used two other restored prairies as comparisons. The first, the Derr West 2002 restoration was planted in the same year as the Derr Pivot, used seed from the same harvest effort, and is located a few hundred yards to the west. It had a more diverse seed mix because it was a more mesic site in alluvial soils, and we created some wetland sloughs that required appropriate seed. Basically, it received the same seed mix as the Derr Pivot site, but with the addition of wetland and wet-mesic species, bringing the species total in the mix to 218. I’ve been collecting data from Derr West every year since 2002, but am only showing data from every other year to match up with the Derr Pivot dataset.
The second site was planted two years earlier, and is called the Dahms 2000 planting. It is adjacent to the Derr West planting (just to the south), so all three of these sites are within the same square mile of land. The Dahms 2000 is also a mostly-mesic site on alluvial sandy loam soils, with some created wetland sloughs. The seed mix for it included 202 species. I collected data every other year from this site, but started doing so in alternating years to the Derr Pivot, so the time scale on the graphs doesn’t match perfectly.
Despite the similar geographic settings of all three, and the same seed mix (mostly) and establishment year for two of them, I expected to see a lot of variation in the rate at which individual species expressed themselves in community assembly. It’s fun to be wrong.
Let’s start with the three forbs I talked about earlier: stiff sunflower, Canada goldenrod, and round-headed bushclover. The two additional sites didn’t have enough abundance of the bush clover to track it, so I eliminated that one from the graphs below. But stiff sunflower and Canada goldenrod patterns were relatively similar between the three sites. They aren’t exact replicas, of course, but there’s more similarity than I’d expected.
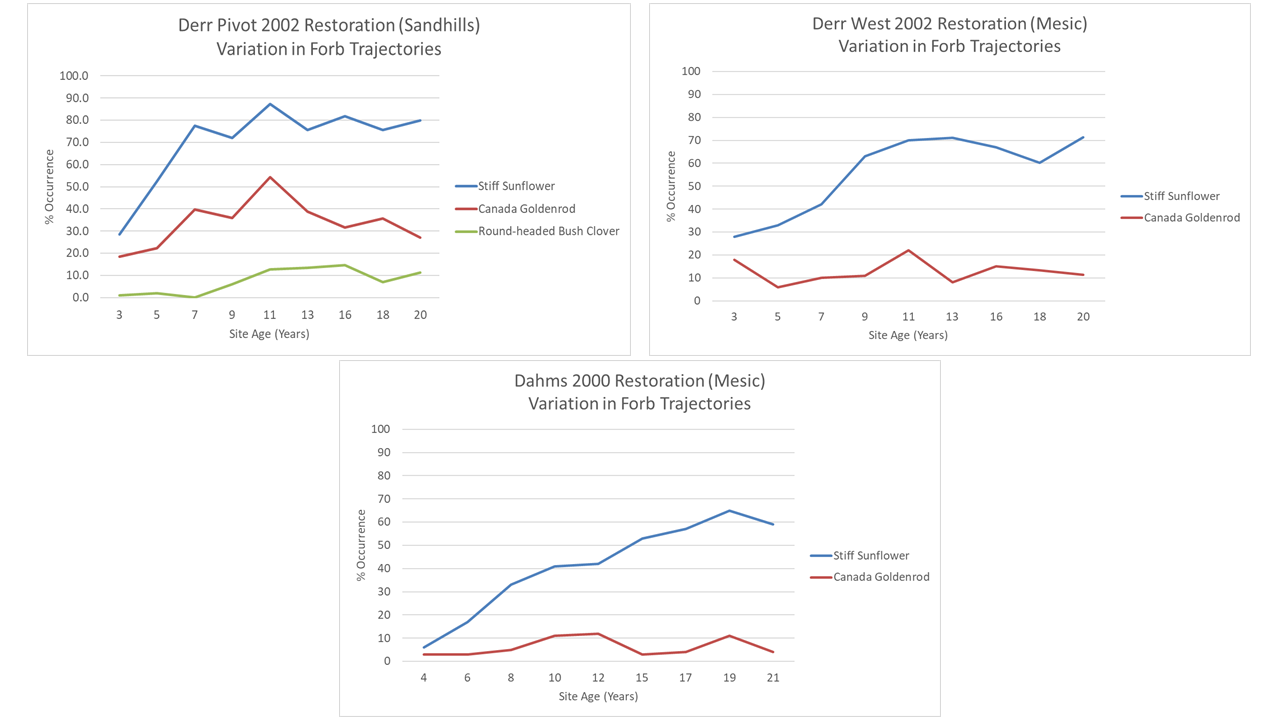
Stiff sunflower took longer to hit its plateau in abundance at the second and third sites, compared to the Derr Pivot, but it still reached similar levels in all three plantings. We had a lot more seed for this species for the 2002 plantings than the 2000 planting, which might account for its faster start in those sites. Either way, though, the species worked its way into about 70-80% frequency of occurrence in all three plantings. Fascinating.
Canada goldenrod is much less abundant at both the additional sites, but in all three sites, its abundance is about the same in year 20 or 21 as it was in year 3 or 4. All three sites also appeared to see a bump in that abundance around 2012, though that may or may not be a real pattern. I know a lot of restoration sites have trouble with Canada goldenrod (or tall/late goldenrod, which in Nebraska is usually treated as a variety of Canada goldenrod). We seem to be fortunate here because Canada goldenrod doesn’t usually act aggressively in our plant communities.
Next, let’s compare smooth brome and Scribner’s panicum, the two grasses that were slowly adding themselves to the community in the Derr Pivot. Scribner’s panicum is in the 2002 site, but at a low enough abundance that it isn’t yet able to be reliably tracked with my data collection method. In the Dahms 2000 restoration, though, it shows a remarkably similar pattern to that in the Derr Pivot, though it waited a little longer to show up in the data. I think we had even less seed for it in the 2000 planting than we did for the 2002 plantings, so that makes sense.
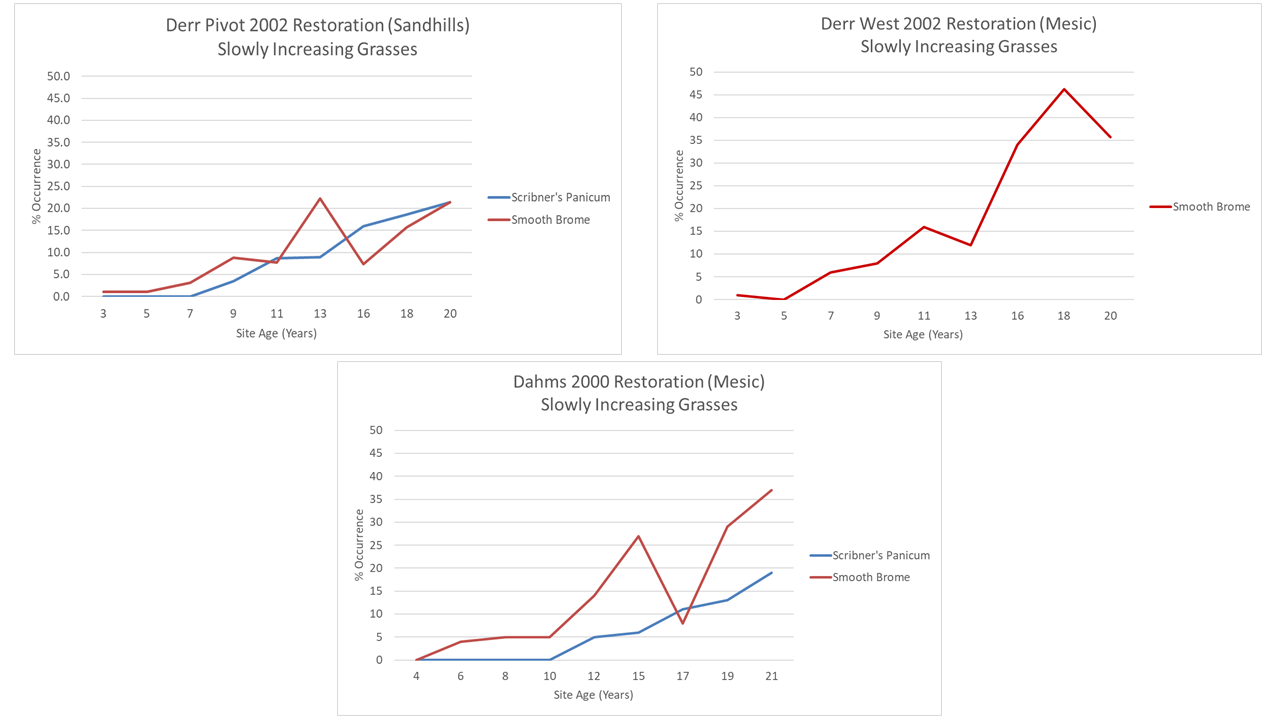
Smooth brome is also showing a very similar pattern of growth between the Derr Pivot and Dahms 2000 sites, including a temporary boost following the 2012 drought. The Derr West site shows a much faster establishment rate, but I can explain that. When that site was still a crop field, there was a weird peninsula of smooth brome and Siberian elm trees that extended into the field. When we prepared the site for restoration, we (I) did a poor job of killing those species out and during wetland construction, and brome sod got spread around the site, speeding its invasion. Sorry about that, everyone.
The similarities continue with big bluestem and Canada wildrye, which exhibit that same opposite pattern of establishment across all three sites. Canada wildrye ended up as a less significant player in the Derr West site than in the other two, but still followed the same kind of path to get there – starting high and the dropping low as the rest of the community filled in around it.
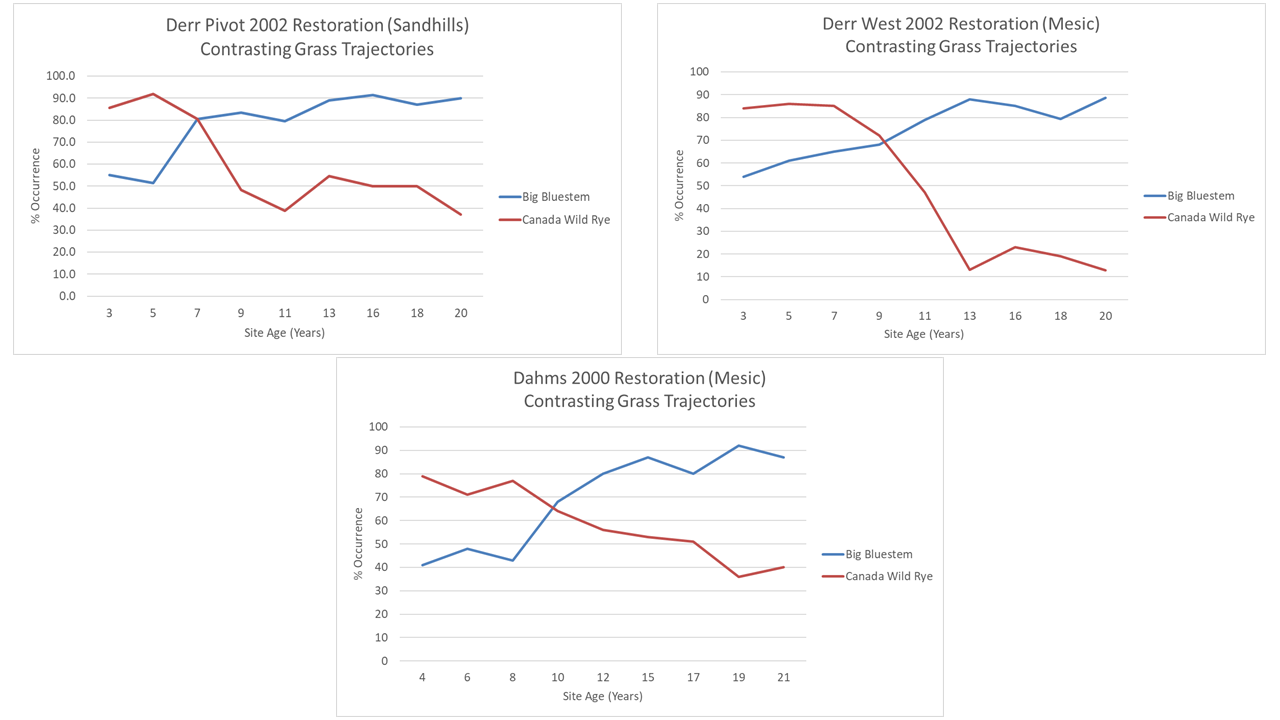
The final graph is the one that most surprised me. I just about fell out of my chair when I first put all three of these next to each other. I’ve spent my whole career being amazed by the complex and (to us) unpredictable nature of prairies and the restoration process. Now I look at these data and wonder if I’ve just been too dumb to notice patterns all around me.

For the record, I haven’t actually changed my mind about the complex and unpredictable nature of prairies. It is really neat, though, to discover some patterns in how things seem to work. I’d love to hear from other restoration sites about whether any data they might have line up with the patterns I’m seeing here. I also wish I’d had time to collect this kind of intensive data from more sites around here to see how consistent these patterns really are.
.
CHAPTER 3 – LAND STEWARDSHIP MATTERS, PEOPLE!
So, the story so far is that three prairies planted in the same area, within a few years of each other, showed remarkably similar patterns of plant community assembly over 20 years or so. All three of those sites have been managed with the same basic stewardship approach that combines prescribed fire and grazing. Patch-burn grazing is probably the best overall description of the approach, but it’s been modified to include periodic full seasons of rest from grazing.

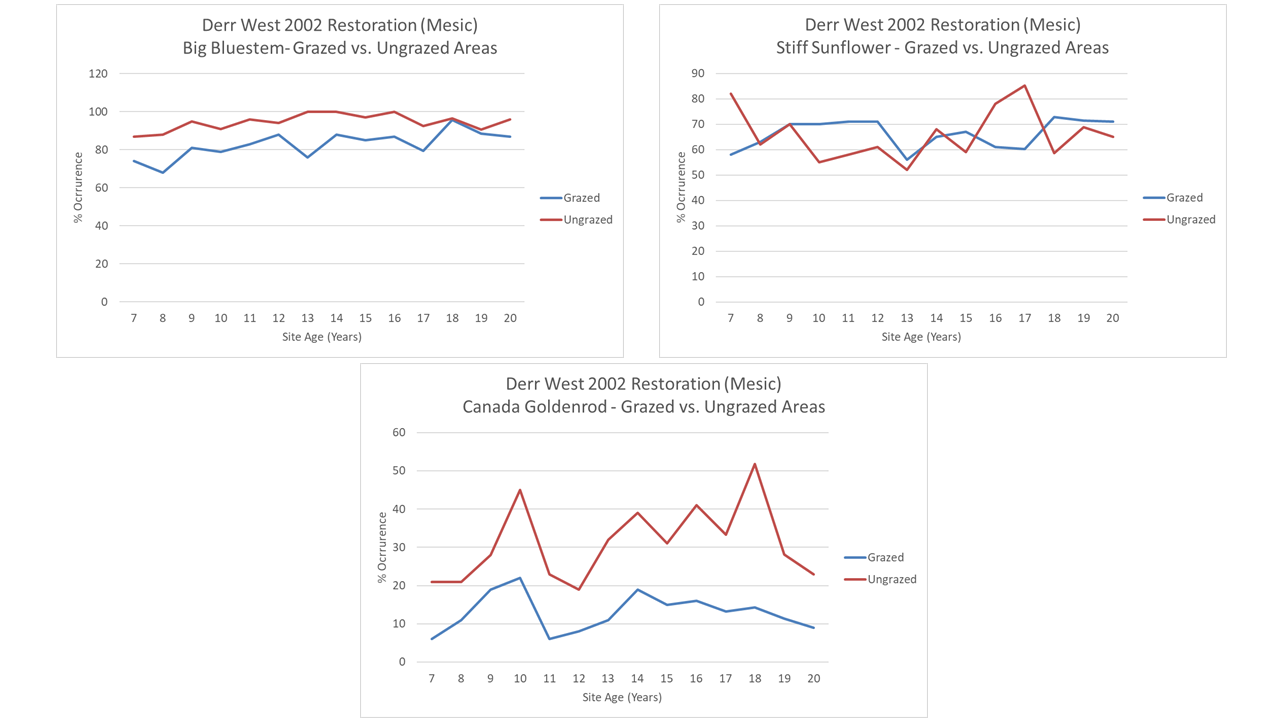
While those three sites have been managed in basically the same way, we did build a nine acre exclosure in the Derr West restoration before we first grazed the site (in year 8 of its life). It receives the same frequency of burning (every 3-5 years) as the surrounding areas but isn’t grazed. It’s far from a perfect test or comparison (it’s not replicated and it’s in the corner of the larger site) but it does allow us a glimpse of how plant species might have established and acted if we were using fire but not grazing.
The following graphs illustrate the response of plants inside and outside of that exclosure at the Derr West restoration. I’m showing annual data here (not every other year) because I have it for this particular site. The first set of graphs below show three plant species for which grazing doesn’t seem to matter one way or the other. Big bluestem and stiff sunflower have maintained very steady patterns of abundance in both the grazed area and the exclosure. Canada goldenrod has a higher abundance in the ungrazed area, but that seems independent of grazing and the periodic ups and downs of its populations are nearly identical inside and outside of the grazing exclosure.
The story changes, though, when we look at the four graminoid species my previous graphs showed are steadily increasing at all three sites. Switchgrass looks like grazing doesn’t affect it much, but Indiangrass, Kentucky bluegrass, and the short-beaked sedge all seem to increase much faster under grazing. The results are pretty dramatic for Indiangrass and Kentucky bluegrass, in particular. If this is representative, the patterns of establishment for these species would have been very different if we’d not been grazing our sites during the later stages of their development. Since these graminoids are major components of the plant community, it’s intriguing to think about the ramifications of that.
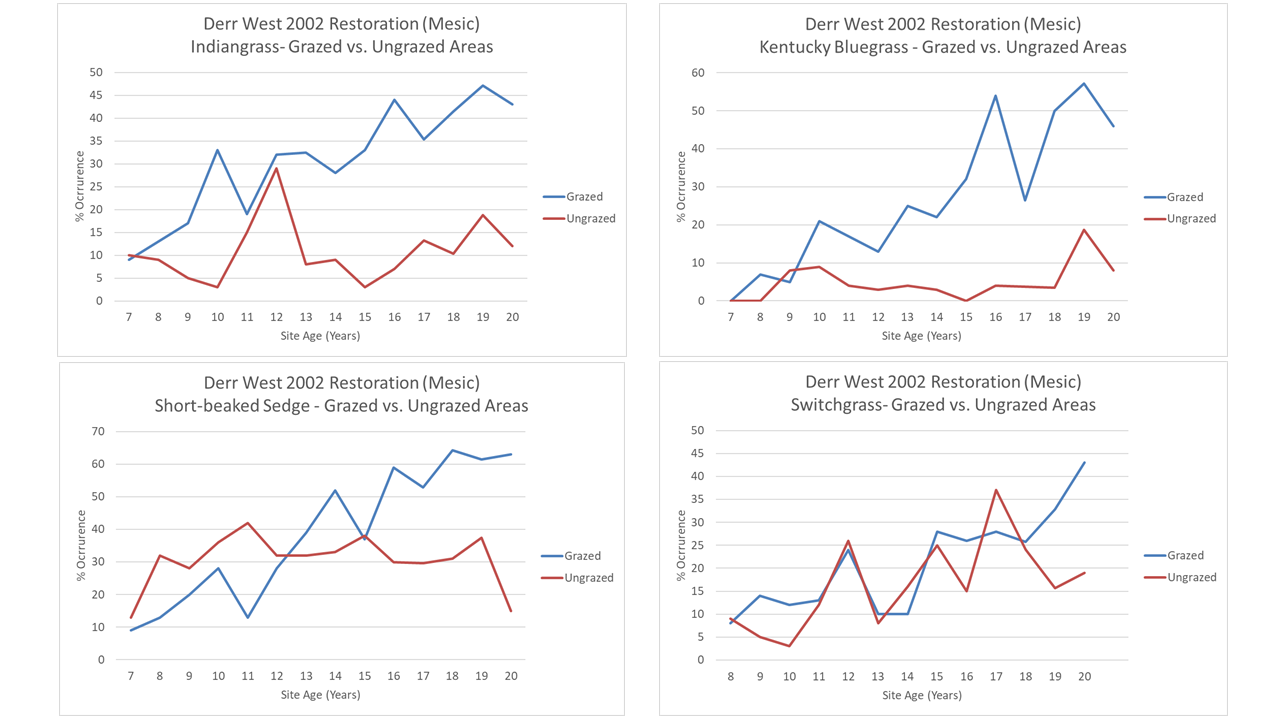
Finally, here are two examples of forbs that seem to show opposite reactions to grazing from each other. Maximilian sunflower (Helianthus maximiliani) has increased over time in the grazing exclosure, but has remained at a steady population level where it is being grazed. In contrast, upright prairie coneflower (Ratibida columnifera) has mostly disappeared from the exclosure, but is still around (more in some years than others) in the grazed part of the site.

Comparing the broader plant communities between the grazed and ungrazed portions of the Derr West restoration is difficult because, again, the single grazing exclosure is in a corner of the larger site and doesn’t have the variation of soils found across the bigger ungrazed area. However, it is interesting to look at the trajectories of each since grazing first started in 2009 (second year of the graphs shown below).
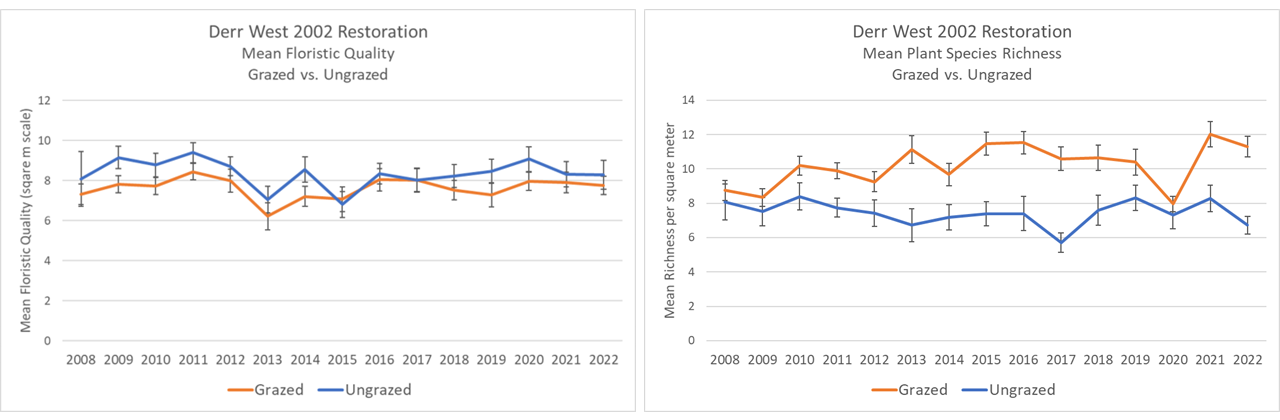
Mean floristic quality hasn’t really changed over time in either site. Mean species richness, however, has increased in the grazed area, gaining an average of 2 species per 1×1 meter plot over time (except in 2020 when excessive thatch suppressed growth). Does that make the grazed site better? Not necessarily, but the difference is intriguing.
Both the grazed and ungrazed areas still have diverse and interesting plant communities, but the management does affect both individual plant species and the broader community composition. In addition, of course, the grazing has big impacts on habitat structure, which is the primary reason we’re using it in the first place. There is a lot more habitat heterogeneity across the grazed site, providing for a more diverse animal community.
So, what can we draw from this long data-heavy post? During the community assembly stages of three restored plant communities, at least many of the major plant species followed remarkably similar trajectories of establishment. In some ways, that should be expected, given the similar sites, seed mixes, and weather patterns. On the other hand, anyone who has spent many years doing restoration work knows how variable results can be from year to year and site to site.
The species establishment trajectories at those three sites, however, seems to have been significantly influenced by management – at least for some species. If we can extrapolate from the grazing exclosure in the Derr West restoration, modified patch-burn grazing increased the establishment rate of some major grasses, but also the overall species richness (at the 1×1 meter scale).

The opportunity to watch plant communities emerge from bare ground and broadcast seed has been one of the most fascinating aspects of my career. We throw one seed mix across a large area and plant species sort themselves into communities that match soil and topography variation. Sometimes, that sorting process matches my expectations, but there are always surprises. Being able to go back and study that process via data helps me understand some of what I watched, but also introduces lots of new questions. I hope I get to watch these same sites for at least another 20 years or so because I don’t think the stories are going to get any less compelling.

THANK YOU! Having less than thirty acres of farm land switched to prairie (Wisc), I really can’t qualify as a nerdy. But no APOLOGIES required on your part! Whether it’s only 1 or 2 percent, of native prairie, still in existence. We need more information on what happens, and what works to create more. Your hope to view those plots in twenty years is exciting. Let’s try to imagine what that 100 acre plot might look like in 500 years?
Harold E
Thanks for this deep dive into this data! I would love to hear about any results you have to share about the efficacy of over-seeding additional species after the initial seeding.
Hi Carolyn, For the most part we haven’t done additional seeding into these restoration plots. There have been a few exceptions and we’ve done some plugging of species that we didn’t have much/any seed for, or that don’t establish well from seed (Carex heliophila, for example, violets, or some wetland sedges). We’ve done more overseeding in some of our degraded remnants, where some of the major forb species disappeared before TNC acquired them. When we do that, we typically incorporate the seeding into our patch-burn grazing, broadcasting seed after a fire and letting the subsequent grazing help control competition during the first year of establishment. That works well for some species, but it takes a lot more seed to get results than when you’re working from bare ground because there’s so much existing competition for those seeds to fight against.
I’m one of the pretty pictures followers and totally read all your delightful commentary! A Nerdy Follower‼️ Many thanks‼️
Sent from my iPad
<
div dir=”ltr”>
<
blockquote type=”cite”>
Great post! Thank you I loved the data. I have a small restoration plot in southern wisconsin and being new to this I learn a lot from your experiences.
How often and time of year did you graze the sites? And for how long?
Generally speaking, the sites had cattle on them most years (4 out of 5 years?) and cattle were present from late April or early May through September or October. Usually, 1/3 of the site was burned in those years, which concentrated most grazing into that portion, but of course cattle wandered through the rest as well. Stocking rates are similar to what NRCS or others would recommend for the site, but the grazing intensity is squeezed into a smaller portion of the site each year, providing the remaining areas with light grazing pressure.
How soon after planting did you start to graze? 8 years?
Most of these sites were grazed at about that 6-8 year mark, yes, but others were grazed a little younger (4-5 years old).
Thank you.
Chris,
Thanks for putting all of this together. I’m definitely looking forward to comparing these patterns with our reconstructions over here (anecdotally, at least).
Restoring cropland to prairie is so rewarding. It’s hard to explain.
But in the name of science, I have wondered of something different.
That is taking cropland out of production and planting seed of every nasty invasive species there is. Let them fight it out and see who wins.
Of course the sight will need be isolated. Perhaps middle of large corn field, and carefully monitored.
I do enjoy all your posts. Keep up your good works.
Andy g.
my money is on canadian golden rod ( although native )
I wonder how Clements and Gleason would interpret these data. Both might find support for their competing theories of plant communities. Adding a few thoughts along these lines would make this fascinating paper even more compelling.
Count me in the nerdy group. Loved it!
Great post. When we established a native grassland through seeding in eastern Pennsylvania, we used four native grass species distributed broadly as a mix in one field, and then we planted the individual species’ seeds as monocultures in areas where we thought they would do best in another field. Naturally, the monocultures started with only the species we planted in each seeding area but, over time, the grasses “sorted themselves out” and invaded one another’s original monocultures as conditioned allowed. The original monocultures are still evident after 20 years, but they’re much more mixed communities than they were in the first few years after planting. Even grasses in the mixed seeding have sorted themselves out according to differences in the field’s aspect, soil moisture, and soil fertility. Given enough time, I think the plant communities in each of these fields are going to grow to resemble one another very closely.
Thanks David,
I’ve seen similar responses to some plots we’ve established as well. What’s most fascinating about that phenomenon, I think, is that plant communities can conform to local conditions as they establish but there are limiting factors. In particular, plants can’t colonize if they aren’t already present in the neighborhood. But it’s even more complicated than that because some plants can come in from seed banks (mostly ‘weedier’ species, in my experience) and others have to come in from the edges. How far they can come varies a lot. In addition, at least here, our colonization process seems to be regulated by precipitation and competition too, so in times of limited precip and no disturbance that suppresses the vigor of established plants, very little colonization happens. When competition for resources is lower (higher precip and/or disturbances) we see more colonization. BUT, even then, there seem to be species that are good at that colonization and others that aren’t. For that latter group, it seems we need to put them in at the beginning (bare ground) of the restoration process if we want them. They seem not to be able to insert themselves into a community that is already full of plants. So much still to learn!