
Melting sea ice might not seem important to those of us living in the middle of a continent. It is.
Weather and climate have always been complicated and difficult to understand, so it’s no wonder that climate change is a topic that confuses most of us. The fact that most climate change discourse is more political than scientific these days makes things worse. It’s hard to have reasonable discussions because most people’s opinions tend to be linked to whichever loud voices they listen to, and few of us understand climate science well enough to draw our own independent conclusions.

The poor groundhog has been a popular scapegoat for this year’s cold spring temperatures. In reality, both this year’s cold spring and last year’s warm spring are much more strongly tied to global warming and melting arctic ice.
I’m certainly not going to wade into the politics of climate change, and I’m not qualified to get very far into climate science. However, I did read something recently that clarified some things for me, so I’m hoping it will help you as well. Thanks to Joel Jorgensen for passing along the article that spawned this post.
One of the most difficult things to understand about global warming is that it can make local temperatures get colder as well as warmer. Here in Nebraska, we’re experiencing a very cold spring – if you can call it spring – this year, but had a very warm spring in 2012. How, you might ask, is it possible that both the warm spring of 2012 and the cold spring of 2013 are a result of global warming?

Last year at this time, pussytoes was starting to bloom in our Platte River Prairies. This year, there’s no indication that they’re anywhere close to that stage.
Scientists have long suggested that more extreme weather patterns (including warm and cold, wet and dry) are a consequence of global warming, but I’ve never had more than a vague understanding of why. Apparently climate scientists are still figuring it out too, but new research published by Jennifer Francis and Stephen Vavrus in Geophysical Research Letters seems to help. After reading a summary of the work in the Omaha World Herald and stumbling through the actual scientific journal article, here is my best shot at explaining the results.
First two pieces of background information you need to understand. This is based on my own rudimentary understanding of this topic, so please take it as such.
1. The warming of the Arctic and the subsequent loss of sea ice is reducing the contrast in temperature between the cold Arctic region and the warmer center of the globe.
2. The contrast between warm and cold areas of the globe is a major driver of weather patterns because it creates an imbalance in atmospheric pressure. The jet stream is the major current of air that tends to run along the boundary between those cold and warm areas (there is actually more than one jet stream, but let’s not get into that). When the jet stream is strong, it moves strongly in a relatively straight west to east direction. However, when it is weak, it makes large north-south loops as it ambles slowly to the east.
Ok, armed with that background knowledge, here’s what’s happening with global warming. Arctic air to the north of us is less cold than it used to be, so there is less contrast between that air and the warm air to our south. That weakens the jet stream, causing it to make large loops as it moves from west to east. Equally importantly, those loops tend to stay in the same place for a long time.
When Nebraska is inside a southward loop of the jet stream, the jet stream’s current allows lots of cold arctic air to come down from the north. That’s what is making our 2013 spring so cold. The opposite is true when we’re inside a northward loop – our weather is dominated by warm air coming up from the south, creating a weather pattern such as the one we saw in 2012. Because a weak jet stream causes those loops to not only be greater in size, but also to stick around longer weather patterns persist for longer periods than they otherwise would. If the weather extra warm for a long time, we tend to have drought, but extended weather periods can just as easily lead to flooding, extended cold temperatures, etc. – depending upon whether we’re north or south of the jet stream current.
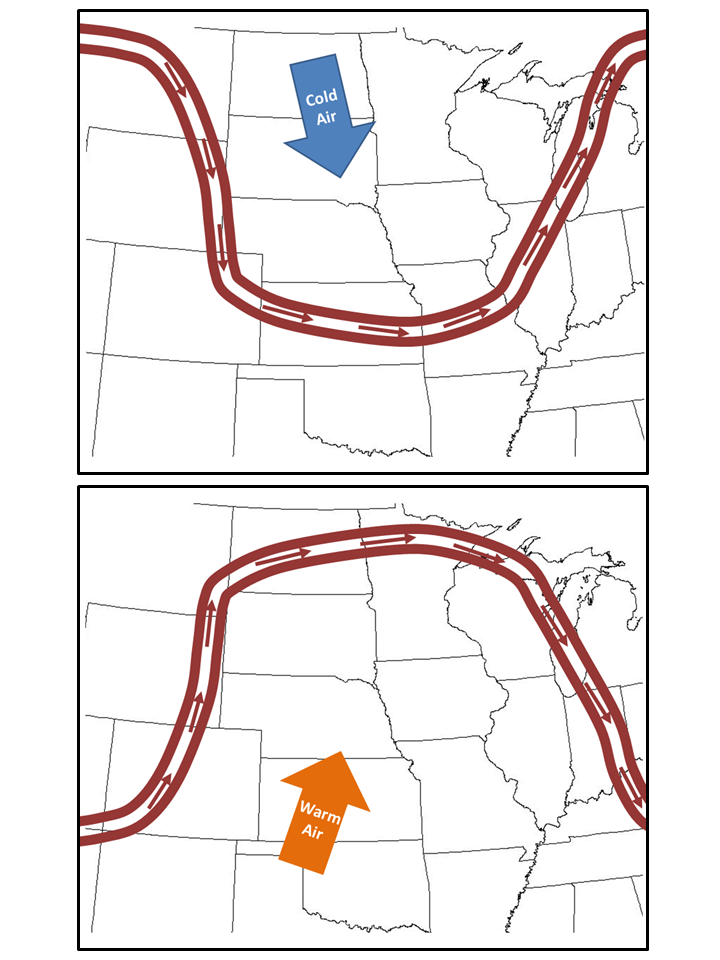
When we are inside a southward loop of the jet stream (top picture) cold air from the north dominates our weather. When we are inside a northward loop of the jet stream (bottom picture) warm air moves in from the south.
Of course, there is much more to weather and climate than just jet stream loops, so a slower, more wandering jet stream is only part of the story. In addition, understanding why we’re getting more extreme and extended weather patterns doesn’t change the situation – it just explains it. I’ve written in the past about some climate change adaptation strategies for those interested in prairie management, restoration, and conservation. A big part of our responsibility is to make prairies as ecologically resilient as possible.
Since creating and sustaining resilience in prairies is largely dependent upon factors we’ve been working on for a long time anyway – species diversity, habitat size and redundancy, etc. – not much changes when we add climate change into the mix, except perhaps that we should feel a little more urgency.
Again, I’m no climate scientist, so I’m trying to explain things I barely understand myself. Please correct me if I’ve mis-stated something or explained things poorly.
")
