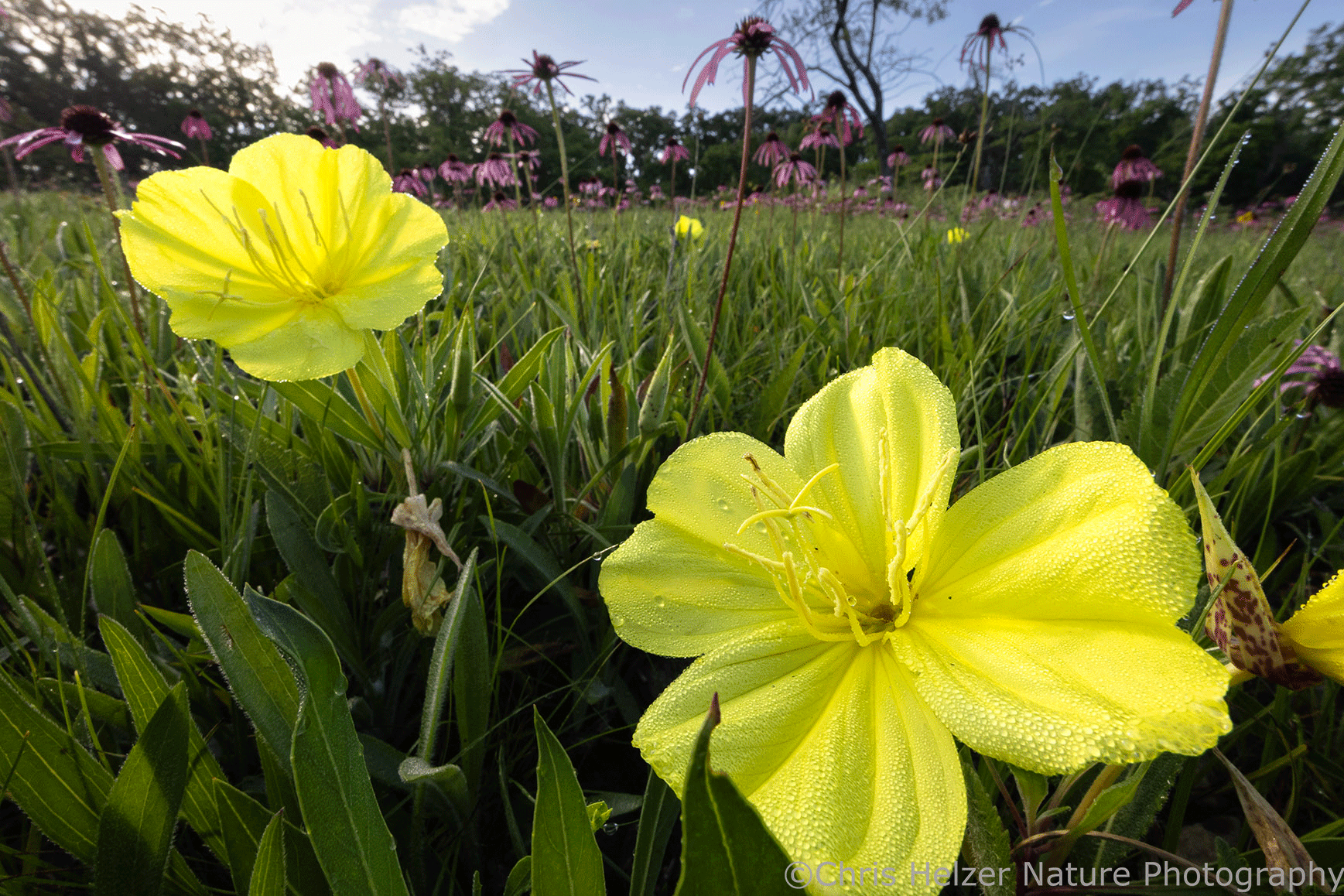Please join us for a fun day in the prairie on Saturday July 11!
Our annual Platte River Prairies Field Day will feature opportunities to learn about the ecology of prairies and wetlands and the identification and natural history of small mammals, plants and invertebrates. You can help with active restoration projects by harvesting seed and make your own art project with pressed wildflowers. If you show up early enough, you can tag along on a photography hike with me. See the full agenda, directions to the site, and other details here.
Master Naturalist Mike Schrad talking about his small mammal research while checking live traps.
This event is located at The Nature Conservancy’s Platte River Prairies and is free and open to all ages. Please use the link above to RSVP so we know how many people are coming and so we can contact if you if we have to change plans because of weather. Bring your own lunch and a refillable water bottle, as well as any other field supplies you need (insect repellant, sunscreen, hat, etc.). You’re welcome to come and go at any time during the day.
Come learn why this is a katydid and not a grasshopper (hint: antennae length) and how to identify the native legume it is sitting on
Here’s one of the top lessons I brought back from the Grassland Restoration Network workshop, hosted this year at the Shaw Nature Reserve in Missouri: There is a convoluted confusion of coneflowers in that part of the world. The prairies I work with in Nebraska have one species of purple coneflower – narrow-leaved purple coneflower (Echinacea angustifolia). It is a wonderful wildflower and more than sufficient for our needs.
Pale purple coneflower (probably) at the Shaw Nature Reserve.
At the Shaw Nature Reserve outside of St. Louis, the story is considerably more complicated. They have pale purple coneflower (Echinacea pallida), of course, which is no surprise in that landscape. However, to add a little whimsy (I assume), sometimes the pale purple flowers can be white instead of pale purple.
I think this is a white version of the pale purple coneflower.
You have to be careful assuming you’re seeing pale purple coneflower, though, because there’s a very similar-looking species found in the many glades at Shaw Nature Reserve. Don’t fret, though – that species, called the glade coneflower (Echinacea simulata) can be easily separated from pale purple coneflower. All you have to do is look at the color of the pollen. If the pollen is white, it’s pale coneflower. If it’s yellow, it’s glade coneflower.
What if there’s no pollen visible? No problem, just say “wow, look at that gorgeous coneflower!”
I believe this is the glade coneflower, and not just because it’s in a glade. I only know that, though, because someone from Shaw Nature Reserve said that’s what it was. See the stump in the foreground? This is a site staff have opened up – removing trees to restore the glade.
To recap, then, they’ve got pale purple coneflower and glade coneflower, and at least one of those can have either pink or white flowers. In addition the pollen of those flowers can be either white or yellow, depending upon the species.
You’d think that would be plenty. Nope. There’s another Echinacea species at Shaw Nature Reserve and this one’s not purple or white. This one’s yellow. A yellow purple coneflower?. That doesn’t seem possible, which is probably why it’s Latin name is Echinacea paradoxa. My wife, Kim, asked me to bring one home with me. I didn’t, but I wanted to.
I’m confident this is the yellow coneflower. I don’t think it matters what color its pollen is.Wow, look at those gorgeous coneflowers!
There were, of course, other species at Shaw Nature Reserve besides coneflowers. Lots of them, in fact. It’s a terrific site with lots of woodland, savanna, and prairie habitat types to explore. We spent a couple days doing just that, along with about 100 prairie restoration practitioners from around the country.
Missouri evening primrose (Oenothera macrocarpa) is an example of another plant species at SNR.Woodland pinkroot (Spigelia marilandica) was another species Kim suggested would look great in our yardPurple milkweed doesn’t grow near me, so it was fun to see a lot of it.They also have animals at SNR, including this gorgeous little five-lined skink (juvenile).
Apart from seeing some new/different plant and animal species, the workshop was also really thought-provoking and inspirational. The staff and volunteers at the Shaw Nature Reserve are doing impressive work to restore and manage prairies, glades, savannas, wetlands, and woodlands (and maybe other communities I’m forgetting). They also do amazing interpretive and educational work with the public. I strongly encourage you to visit the site.
We talked about some specific land stewardship/restoration tactics, including the use of grass-specific herbicides to suppress grass dominance, especially when paired with overseeding of forbs. SNR staff also showed us a lot of tree thinning/clearing projects and talked about what appear to be successful strategies. In addition, there were lots of side conversations about other invasive species and restoration lessons. Many of the stories we talked about can be found at the Grassland Restoration Network blog site, which, if you’re not following it, you should: https://grasslandrestorationnetwork.org/
I also encourage you to check out this storymap, which showcases a big project SNR staff have recently worked on. It’s coming along really nicely. Not only is the project impressive, it’s one of the best applications of the storymap format I’ve ever seen.
Learning togetherSome of the beautiful landscape at SNRSome kind of EchinaceaA close-up photo of some kind of Echinacea. It’s likely the same species as the one just above it. Or maybe not. There’s no way to tell, as far as I know.I know this one! This is goat’s rue (Tephrosia virginiana), another of the countless plant species at SNR I don’t see in my local prairies.Prairie dock (Silphium terebinthinaceum)An ambush bug hunting on wild quinineAn attractive little damselfly
Huge thanks to the Shaw Nature Reserve staff and volunteers for their incredible work and for hosting a terrific workshop for us. Next year’s workshop will be in Nebraska – co-hosted by Prairie Plains Resource Institute and The Nature Conservancy. Stay tuned for more details.